


(本稿は「農業および園芸」2012年新年号および2月号に掲載予定記事を一部改変したものである)
1.はじめに
2011年3月11日午後2時46分に発生した東日本大震災は,未曽有の規模の地震とそれに続く津波により東日本太平洋沿岸地域に甚大な被害をもたらした.農業生産においても農地,施設の破損,海水の浸入など重大な影響を受け,津波による流失及び冠水等の被害を受けた水田面積は約20,151haで,農地全体の約2.6%にものぼると推定されている(農林水産省 2011年3月29日現在).冠水した水田では多くの関係者の努力により除塩作業が進められている.しかし灌漑・排水システムの損壊や地盤沈下により速やかな除塩が困難な場合もみられる.また,利根川流域など一部地域では液状化による噴砂の堆積や圃場の損壊が発生したが,ここでも噴砂にともなって高塩類濃度の地下水が土壌や水路へ混入し問題となっている.これらの地域での作物生産の早期の再開には,塩害を軽減する耐塩性作物,品種や栽培法の利用が有効と思われる.本稿では,水稲を中心とした作物の塩害の生理機構と栽培,品種対策について紹介する.
2.塩害の種類
(1)海水の特徴
農作物の塩害は,海水の影響を受ける海岸部や塩類の集積しやすい半乾燥地,岩塩地層をもつ地帯を中心に広く世界でみられ,その克服は農業上の大きな課題である.日本では津波,高潮,潮風害に伴う海水の農地への侵入による塩害の他に,降水が直接かからず施肥量が多い施設園芸土壌でも塩類の過剰集積による生育障害が見られる場合がある.海水や岩塩が由来の塩類の場合は多量のNaClを含む.一方半乾燥地土壌では一般にCa,Mg,Naの塩化物,硫酸塩および炭酸塩を多量に含む.施設園芸の塩類障害土壌では肥料に由来するCa,K,Mgの硫酸塩,硝酸塩など多様な生理活性の強い成分を含むことが多いのが特徴である.
海水の塩類濃度・組成は海洋の場所や深さ,季節などにより変化するが,平均で約3.4%程度の多量の塩分を含む.海水組成はカチオンではNa,アニオンではClの濃度が著しく高く,またSO4も高いのが特徴である.海水でNaが高いのはその生成過程においてNaがCaやMgにくらべて炭酸塩として沈殿しにくかったことによると考えられる.また高いClは火山ガス由来と考えられている.1価カチオンのNa/K比が海水では27.6程度と河川水(2.74)や土壌(1.13)より著しく高い.高等植物ではKは必須要素であり,Naより高く維持される必要がある.このため海水にさらされた場合,イオン吸収の制御が必要となる.また2価カチオンではMgがCaよりも高く,河川水ではCaがMgより高いことと異なる.
(2)塩類土壌
一般に塩類濃度の指標として電気伝導度(EC)を用いることが多い.ECは,測定が容易であり,また溶存イオンの総濃度(活動度)を反映すること,作物の吸水に影響する浸透圧とほぼ比例すること,などから作物への影響程度の指標としてよく用いられている.土壌のECは土壌飽和溶液や懸濁液で測定される.土壌サンプルが硫酸カルシウムを含まないときは,土壌飽和溶液のEC(ECe)と懸濁液で測定したECとは次のように変換できる(IRRI);
ECe=2.2 × EC1:1 EC1:1は土壌と水が1:1の懸濁水での測定値
ECe=6.4 × EC1:5 EC1:5は土壌と水が1:5の懸濁水での測定値
またEC値を用いて,浸透ポテンシャルが推定できる.
浸透ポテンシャル(MPa)=EC(dS/m)× 0.036
ただし,これらの関係はあくまで目安であり,塩類の組成,土壌の性質により変動する.圃場容水量以下で栽培される畑作物においては,実際に根が接する土壌溶液のECはECeより高くなる.
USDAの基準では塩類土壌はECeと交換性Naの割合(ESP;Exchangeable Na percentage, ESP=交換性Na/CEC x 100),pHを組み合わせて以下のように分類されている(US soil survey staff 1951).
| 塩性土壌(saline soil); | ECe>4,ESP<15,pH<8.5 |
| アルカリ土壌(alkaline soil); | ECe<4,ESP>15,pH>8.5 |
| アルカリ-塩性土壌(saline-alkaline soil); | ECe>4,ESP>15,pH>8.5 |
(3)海底堆積物の影響
津波は海水に加えて農地表層へ海底泥土の蓄積を伴う場合がある.海底泥土は,多量の塩分や重金属を含むその他の無機物や有機物を土壌に蓄積させるだけでなく,透水を妨げ,降雨や灌水による除塩の能率を低下させる.津波によって運搬された泥土は硫化物を含む場合があり,これらが酸化されると硫酸酸性を引き起こし,作物生産上の問題となる.硫酸酸性による障害機構は1930年代に霞ケ浦や各地の干拓地で初めて解明された.潮間帯などでは,海水中の硫酸イオンと堆積物中の酸化鉄が低酸素条件下で還元,反応し硫化鉄(FeS)やパイライト=黄鉄鉱(FeS2)が生成する.硫化鉄は陸上で酸素が供給されるとSO42-を生じて土壌を酸性化し,作物生育を阻害する原因となる.
FeS+(15/4)O2+(7/2)H2O=Fe(OH)3+2H2SO4
土壌酸性は畑では特に問題となる.一方,水田ではSO42-は湛水条件下で還元され,硫化水素になる.多くの硫化水素は土壌中の鉄と難溶性の硫化鉄(FeS)になり無害化される.しかし,鉄含量の少ない土壌では硫化水素や硫化物イオンが遊離し,水稲根の養分吸収阻害や根腐れの問題を引き起こす.
3.津波・高潮による農地被害
過去にも国内外で多くの津波や高潮による農地被害の事例があり,対策がとられてきている.
(1)国内の事例
1959年9月26日に紀伊半島から東海地方に襲来した伊勢湾台風は三重,愛知,岐阜の3県の農地に甚大な被害を与えた.各所で高潮により堤防が決壊し,農地への冠水・浸水,土壌の流出などによる被害面積は水田を中心に約32,000haに及んだ.愛知県における詳細な調査(愛知県 1963)によると浸水期間は長い場合には2ヶ月~4ヶ月におよび,多量の塩分が蓄積した.交換性カチオンではNaとMgの割合が高く海水の影響が顕著であった.また流入泥土層を形成した場合には,多量のCl-とSO42-が混入していた.次年度の作付けでは,Cl-が0.08%(乾土当たり)以上,SO42-が40 mg/100g(風乾土当たり)以上の水田で,水稲の生育や収量に影響がみられている.
1960年5月23日に発生したチリ沖地震により,翌24日から三陸海岸を中心に津波が襲った.宮城県では水田683ha,畑253haが冠水被害を受けた(宮城県立農業試験場1961).排水良好な圃場では塩類濃度は迅速に低下したが,排水不良地帯では塩害により水稲,麦,蔬菜に影響があった.開花受精中に冠水をうけた麦は収穫皆無となった.水稲では9月に土壌塩類濃度が0.1-0.3%(乾土)の圃場では20-80%の減収がみられた.ダイズではCaや堆肥施用による生育促進効果が報告されている.
その他近年では,1999年9月24日の台風18号における熊本県での高潮,2005年8月30日に香川県を来襲した台風16号にともなう,海岸部の高潮や河川での海水の逆流,2006年10月7日の福島県相馬市での高潮,平成2009年2月24日の富山県東部での暴風による高潮などの事例がある.
(2)インドネシア沖地震
2004年12月26日に発生したスマトラ島西方沖地震(マグニチュード9.1~9.3)によって,インド洋に面するインドネシア,タイからインド,東アフリカまでの広い海岸域を津波が襲い,農業活動にも甚大な被害をもたらした.このインド洋津波は農地に多量の塩分と堆積物をもたらした.アンダマン諸島での土壌のモニタリングによれば,降雨が十分あり(年間3000mm以上),排水条件が良好であれば,2年以内で土壌塩類濃度は大きく低下した(Rajaら 2009).降水量の多いインド南部でも2年以内に農地が回復した例が多い(Chandrasekharanら 2008,Kume ら 2009).ただし,地下水の塩類濃度の低下は遅いことや,海岸部から離れた地域での地下水のくみ上げが海水の侵入をもたらすことへの留意の必要性も指摘されている.一方で,排水システムが破損した場合は,3年間で3000 mm以上の積算降水量があっても除塩が充分に進まなかった場合もある.タイでは排水溝により除塩促進効果を上げている(Nakaya ら2010).またゴム園での観察では,2005年の雨季初期の7月までに一度樹液量は回復したが,雨季終了時の11月に再び品質は低下したという(中矢ら2006).おそらく降雨の影響とみられるが,多年生作物では障害の長期的な変動にも留意が必要と思われる.
4.塩害の生理メカニズム
土壌の高塩類濃度により植物の生育が阻害される生理メカニズムは,大きくわけて浸透圧ストレスとイオンストレスからなる.浸透圧ストレスは,根圏の水ポテンシャルの低下により吸水が阻害され,植物の水ポテンシャルの低下,気孔の閉鎖,光合成の低下や葉の伸長抑制を引き起こすことで生育を阻害する.また葉内ではアポプラストのNa濃度が水耕液の10倍以上にも高まり,葉細胞の脱水を引き起こす(Flowers ら 1991).イオンストレスは,植物体内に入った過剰なNaClイオンが代謝を阻害し,葉の枯死や生育の阻害をもたらす.過剰なNaによる他のイオンとのアンバランスも生育障害の原因となる.
浸透圧ストレスとイオンストレスは,多くの場合複合的に作用するため,それぞれの関与を明確に分離することは困難である.一般に浸透圧ストレスは,塩処理の初期より影響し,より長期間の処理では体内への塩類の集積が進み,イオンストレスの影響が大きくなる(Munns and Tester 2008).また部位別にみると,成熟葉では塩類の蓄積によるイオンストレスが卓越し,新しい葉では水ストレスの影響が大きいと考えられる.
生育に大きく影響する光合成能は,浸透圧ストレスによる気孔閉鎖やイオンストレスによる機能障害により低下する.特に成熟葉ではNa濃度が高まるため,光合成の抑制程度が大きい(Yeo ら 1985).塩処理下では光合成の低下にともない発生する活性酸素の消去系の活性が高まる.また塩処理により根の呼吸速度が増加するが,これは浸透圧調節やイオンの選択吸収,膜の修復などにエネルギー消費が増えるためであろう.
イネではNaClと浸透圧を調節するためのポリエチレングリコール(PEG)による生育抑制程度を同等の浸透圧で比較すると,NaClの場合50mMですでに生育抑制がみられるが,PEGのみでは100mMまで抑制は小さいことより,少なくともこの濃度域ではイオンストレスの影響が大きいことが示唆される(Matoh ら 1986).またNaClのみによる生育障害は,総塩類濃度を同じにした海水を与えた場合よりも大きいことより,イオン間のアンバランスも生育抑制を大きくすると考えらえる.特定のイオンが高いと他のイオンの吸収や代謝における機能を抑制しやすい.NaはK,Mg,Ca の吸収を,またMgはK,Caの吸収を抑制しやすい.NaClによるMn吸収抑制,Pの吸収,移行の抑制もみられる.特にNaが大量に存在する場合には,多くの酵素反応に必要なKとのバランスを積極的に安定化する必要がある.
イオンストレスの作用には,細胞質中のイオンによる酵素の阻害,活性酸素の増加,脂質の酸化などがある.海水中のイオン種の作用程度は多様であり,また栽培環境,作物種,ステージにより異なる.NaとClの作用の違いについては必ずしも生理メカニズムは明確ではないが,作物間で異なる可能性がある.イネ(山内ら1987)やコムギ(Gorham et al. 1990)などイネ科作物では,地上部の生育阻害は地上部のCl濃度よりもNa濃度とより密接な関係を示すことから,Naの影響が強いようである.ラッカセイ,インゲン,ダイズなどマメ科作物などでは一般に葉のCl濃度がNa濃度より高く影響が大きいと思われる.ClはSO4より生育や光合成の抑制作用が大きい場合もある(Marschner 1995).ブドウ,レモンなどでも相対的にNaよりClの体内集積が多い.土壌の過剰なMgは,キュウリでは根のミトコンドリア機能の障害などを引き起こし,地上部生育を抑制した(嶋田1972).
実際の水田や畑地における塩害では,土壌のpHの変動,微生物活性の低下,還元の促進,ホウ素など微量要素の過剰障害,など多様な要因がイネの生育に影響を及ぼすため,そのメカニズムは複雑である.高pH下ではZn欠乏を,硫酸酸性土壌のような低pH下ではFe過剰を引き起こす.加えて,塩害は土壌物理性を通しても作物生育に影響する.土壌粘土はマイナスの電荷を持ち,カチオンを吸着している.通常の農地土壌では,Ca2+が多くを占めるが,海水に含まれるNa+やMg2+と土壌に吸着したイオンとの間で交換反応が起こり,土壌の吸着イオンとしてNa+とMg2+の割合が高くなる.Na+が多く吸着した粘土は,水に分散しやすくなり,土壌の間隙に詰まり,透水性を悪化させる場合がある.これにより,畑作物の根の酸素不足や,除塩の効率低下などの問題を引き起こす.
5.耐塩性の作物間差異と耐性メカニズム
植物の耐塩性メカニズムとしては大きく浸透圧ストレスとイオンストレスに対するメカニズムがあり,関与する候補遺伝子群に関する知見も集積されつつある(表1).浸透圧ストレスを軽減するメカニズムには浸透圧調節,水吸収の促進,蒸散の抑制などがある.イオンストレスに対しては,大きくは,体内特に地上部へのNaの移動・蓄積を低減する回避(Na排除)と,Naを体内組織内に取り込んだ場合の組織耐性の2つのタイプに分けられる.前者には根や葉などからのNaの体外への排出,地上部から根への還流,組織の水含量の増加による希釈などがある.また後者には,液胞へのNaの隔離,カウンターイオン溶質の合成,KによるNaの置換などがある.高等植物では酵素そのものの耐塩性には大きな差は認められておらず,何らかの手段で酵素活性をNaから保護している.耐塩性を付与するこれらの過程に関与する分子機構もすこしずつ明らかになってきている.根が高塩環境にさらされると短時間で地上部に反応がみられることから,根から地上部への何らかのシグナル伝達機構があると考えられる.初期の根の反応では細胞質Caの増加がみられ,SOS3の関与が示唆されている.Naの体内輸送や体内K/Naの維持には,高親和性KトランスポーターHKTファミリーが関与している.耐塩性の品種間差異にHKTファミリーが関与する可能性が,コムギ(James ら 2006a,Genc ら2010)やイネ(Ren ら 2005)で示唆されている.
高等植物は大きく高塩環境に適応した塩生植物halophyteと適応性が低い非塩生植物glycophyteに分けられるが,多くの作物はサトウダイコンなど一部を除けばほとんどが中生植物である.乾燥地や海岸沿いの地帯に多く生息する塩生植物の多くでは,体内にNaClを高濃度に蓄積しても細胞機能を維持できる耐性メカニズムをもつ(高橋1987).また多肉化により蒸散を抑えている.海岸域に生育するホソバノハマアカザなどでは高濃度のNaを体内に蓄積するが,液胞内に隔離することにより細胞質の機能障害を回避している.液胞ではClやシュウ酸でNaとのイオンバランスをとっている.一方浸透圧調節のために細胞質ではショ糖やアミノ酸(ベタイン,プロリンなど)を集積する.河原などにみられるヨシは,塩生植物ではないが広い塩濃度で生育できる.ヨシでは地上部Naが過剰にならないようにNa吸収を制御するとともに,地上部から根へ師管を通してエネルギーを使って再輸送し排出する機能が高く,Naの地上部への移行率を低く抑えていると思われる(Matsushita and Matoh 1992).
作物間でも耐塩性に差異があり,生育や収量に影響する塩類濃度の閾値には大きな幅がある.アカザ科で好塩性作物のサトウダイコンは,100mMまで生育抑制はなく,NaをK以上に体内に蓄積できる.これはNaをKに代替して代謝に利用できるためと考えられ,Na施用による生育促進効果もみられる.イネ科ではオオムギで耐性が高く,イネは低~中程度とされている場合が多い.オオムギの耐性品種では少なくとも葉内Na濃度が400mM程度まで光合成活性を維持することができるが,これにはNaの液胞への隔離と浸透圧調節機能が寄与している(James ら 2006b).マメ科作物においては,根粒の窒素固定活性が塩類ストレスへの感受性が高く,また宿主の耐塩性が弱い場合に特に窒素固定活性は抑制をうけやすい(池田ら 1987).
作物間・品種間の耐塩性の程度と体内へのNa蓄積の関係は単純ではないが,一般に耐性の高い作物種では体内Na濃度が高くても感受性の作物より生育の抑制が小さい.また,同一作物内で比較するとNa濃度が低い品種で耐性が高い傾向があり,イネなど耐性が比較的低い作物種で特にこの傾向が強い(山内ら1987など).コムギでの品種間差異もNa排除に関連していた(Munns and James 2003).一方で耐塩性の高いワタの品種間では耐性の程度とNaの葉への蓄積との関係は一定ではなく,耐塩性の品種でNaが高い事例もみられる(Leidi and Saiz 1997).オオムギやトマトでもNa濃度と関係がみられなかった場合もある(Dasgan ら 2002).
{kind=link}
6.耐塩性のイネ品種間差異と有用特性
イネでは品種間で耐塩性に変異があることが知られている.例えば高塩類条件下(NaCl 80mM)での対照区に対する地上部乾物重で23-80%の変動がみられている(山内ら1987).イネでの品種間比較の評価指標としては,乾物生産や収量の他に,苗の生存率,葉の枯死・萎れ程度,体内Na/K,体内Na濃度などが用いられている.クロロフィル蛍光をもちいた光合成への影響程度も利用されている.1970年代より国内外で幅広い遺伝資源を用いたスクリーニングが行われ(Akbar ら 1972,Datta 1972,など),インドの沿岸部の在来インド型品種である「Pokkali」や「Nona Bokra」など高い耐塩性をもつ品種が見出された. イネ品種間の耐性の違いには,浸透圧ストレスおよびイオンストレスへの耐性が関与していると考えられるが,長期間のストレス条件では主に後者の影響が強いと思われ,根のNa吸収抑制,新葉への移行の抑制,葉中高濃度Naへの耐性などが重要な形質とされている(Yeo et al. 1990).一般に耐性品種では葉,特に上位葉のNa濃度やNa/Kを低く維持する場合が多い(Flowers and Yeo 1981,Yeo and Flowers 1986,Akita and Cabuslay 1990など).これは根でのNa排除能が高く,また根,茎や下位葉に多くNaを蓄積する能力が高いことによる.また特に初期生育の耐性が高い品種は高い生育量をもつ傾向がある(Yeo ら 1990)が,Na濃度を高めない形質として有効と思われる.さらにPokkaliの耐性には液胞へのNaの隔離,アポプラストNa濃度を低く維持する能力など,組織間でのNa分配能力も関与すると考えられている(Anil ら 2005).プロリン集積能など浸透圧調整能も寄与すると想定されるが,Nona Bokraでプロリンの集積は特に高くはなかったとする報告もある(Lutts et al 1996).また,耐性品種では塩類ストレスに敏感に反応して気孔を早く閉じたり,活性酸素消去機能が高い傾向がみられている(Moradi and Ismail 2007).このように耐性品種では複数のメカニズムが耐性を付与していると考えられる.
7.分子機構と今後の育種方向
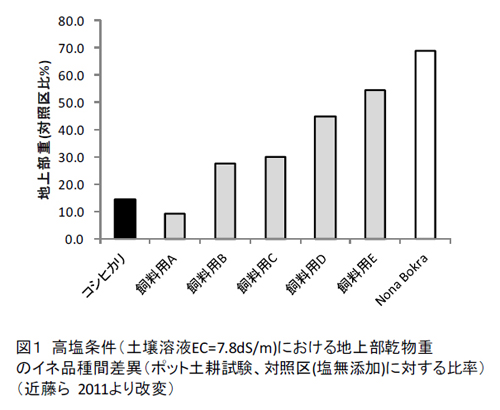
PokkaliやNona Bokraを母本としてIRRIを中心に熱帯での栽培適性のある耐塩性品種が育成されている.葯培養によっても耐性品種Bicolが育成されている(Senadhira et al. 2002).一方,日本では耐塩性品種の必要性が低かったことがあり,品種育成の事例は限られる.通常の食用品種には,塩害に強い耐性を示すものはこれまでほとんど見出されていない.農研機構・九州沖縄農業研究センターでの飼料用品種を用いた試験では,インド型飼料用品種「モーれつ」が生育障害の程度が軽いことが示されている(加藤ら 2006).最近の実験では,その他の複数の飼料稲品種にも比較的耐性のある品種の存在が示唆されてきている(近藤ら2011).ポット試験では,これらの品種ではNonaBokraほど耐性は高くないが,高塩類条件(土壌溶液EC7.8dS/m)での乾物生産がコシヒカリを上回り対照区比で最大50%以上の乾物重を維持していた(図1).ただし,籾生産は抑制を受けやすいため,茎葉部利用が望ましい.今後より幅広い実用品種の耐塩性評価とその活用法の確立が待たれる.
耐塩性品種育成を加速するためには分子機構に基づいた有用な量的遺伝子座(QTL)の集積が期待される.耐性のPokkaliと感受性のIR29ではAKT-1タイプKチャンネルの変動に違いがみられることや(Golldack ら 2003),栄養生長期や幼穂形成期において耐性品種・系統では感受性品種・系統より塩処理下で変動する遺伝子群が少ないこと(Walia ら 2005,2007)など,分子レベルでの品種間の反応の違いも明らかにされつつある.塩処理下で耐性品種・系統で遺伝子発現の変動が小さいことは,葉内Na濃度の安定性を反映している可能性があると思われる.
PokkaliやNona Bokraについては,その耐塩性にかかわるQTLの同定や機能解析も進展している.耐性の高いNona Bokraと感受性のコシヒカリの交配集団を用いたQTL解析(Linら 2004)では,幼苗を140mMのNaClを含んだ水耕液下で生育させ,生存日数,根および地上部のNa+,K+の濃度を指標として解析をしている.その結果,複数のQTLが検出され,そのうち第1染色体に検出したQTL SKC1は,遺伝子型がNona Bokra型で地上部のK+の濃度を高める効果を示した.さらに,第7染色体上に,遺伝子型がNona Bokra型で地上部のNa+濃度を下げる効果を有するQTL SNC-7を検出し,その寄与率も48.5%と高かった.このうちSKC1遺伝子はNaトランスポーターであり,地上部のK,Na濃度を制御することにより耐塩性を高めていることが示唆されている(Ren ら 2005).このトランスポーターは地上部,根の導管周辺で発現が高く,Naイオンを地上部から根に再循環させて,Naの根からの排除を促進すると推定されている.
Pokkaliと感受性IR29の交配集団でも複数のQTLが見出されており,このうちNa-Kを制御するQTLSaltolについては詳細な解析が進んでいる(Thomson ら 2010).この領域はPokkali型でNa/Kを安定化させる作用があり,SKC1と同じである可能性が高いこと,複数の遺伝子を含む可能性があること,が示唆されている.同時にこの領域単独では塩処理下での葉の枯死程度でみた耐塩性の向上効果は大きくないことも示されており,十分な耐性の付与にはQTLの集積が必要と思われる.このようにイネ品種の耐塩性には複数の生理メカニズムと遺伝要因が関与している.また苗期,栄養成長期,生殖成長期などステージ間により品種間差異が異なる場合もみられている.それぞれのステージに有用な形質を組み合わせることで総合的な耐塩性の向上が図られる.塩処理が長期化し,体内Na濃度が高まるのにしたがい,浸透圧ストレスへの適応性やNa排除能の寄与よりもNa耐性の寄与が大きくなるものと想定される.
PokkaliやNona Bokraなどインディカの他に,ジャポニカ遺伝資源にもNa/Kの維持に関与する遺伝要因が存在することも示唆されており(Ahmadi ら 2011),新たなQTLの活用も期待される.
{kind=link}
8.栽培技術対策
(1)土壌塩類濃度の指標
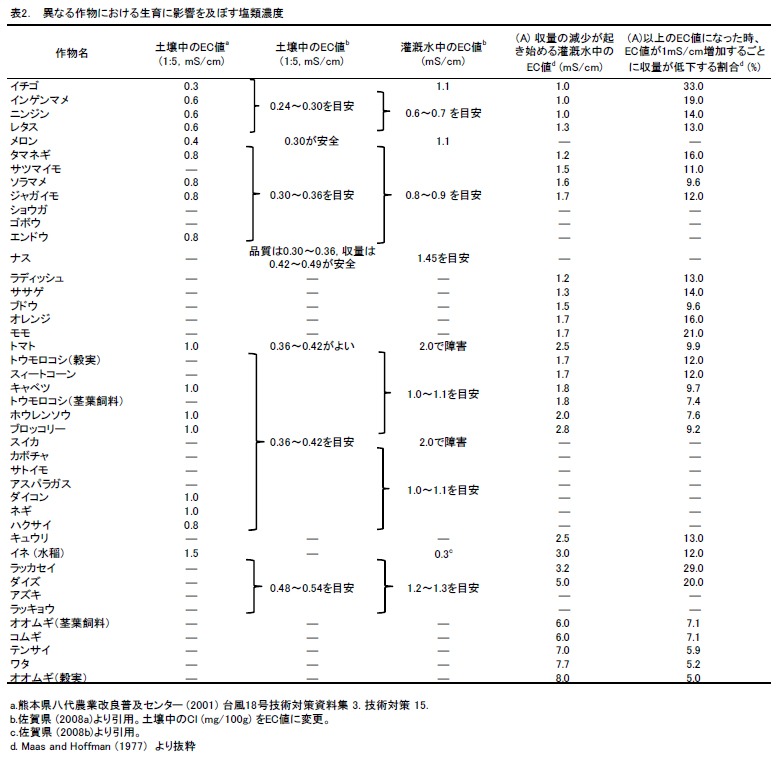
高塩類土壌での作物栽培にあたっては,土壌条件にあわせた作物・品種の選択が有効である.生育や収量に影響する塩類濃度の閾値には作物間で大きな幅がある(表2).ただし,このような閾値には栽培環境,品種などにより変動があることには留意が必要である.また生育ステージよっても耐性は異なる.水稲では,活着期,穂ばらみ期および出穂開花期に特に塩類ストレスに感受性が高く収量への影響が大きいとされる.日本における水稲の除塩目標値はおおむねEC値(1:5抽出)0.3 dS/m~0.7 dS/mとされている場合が多い.葉齢2.0のコシヒカリの苗を用いた場合,土壌塩分0.1%(EC値(1:5抽出)で0.3 dS/m)で活着に影響がみられ,土壌塩分0.15%以下で苗の活着程度80%が確保されている(木田ら 2007).IRRIでは水稲の塩害の指標として,土壌飽和抽出液の電気伝導率(ECe)や交換性Naパーセント(ESP)と減収の関係を,おおよそ次のように整理している(IRRI).
・ECe<2 dS m-1 最適条件であり,減収は認められない
・ECe>4 dS m-1 わずかな減収(10-15%)が認められる
・ECe>6 dS m-1 20-50%の減収が認められる
・ECe>10 dS m-1 耐塩性を有しない品種では50%以上の減収が認められる.
{kind=link}
ESPについては;
・ESP<20% 著しい減収は認められない
・ESP>20-40% 10%程度の減収が認められる
・ESP>80% 50%減収となる
土壌の限界EC値は,塩類組成,塩類が蓄積した土壌層位,生育ステージなどの違いで変動する.強風や高温の場合には蒸散が高まり,塩類障害が助長されやすいなど気象条件も影響する.また,同じ作物中でも品種によって耐塩性に大きな幅があるため,実際の栽培にあたっては注意が必要である.
圃場の除塩方法としては,淡水を灌漑し,暗渠を利用して排水する方法と,代掻き・落水により地表排水する方法がある.前者で効率が高い場合が多い.いずれの場合にも塩類濃度の低い灌漑水が必要であり,適切な排水が行えることが迅速な除塩の条件となる.実際の脱塩・除塩法については各種資料(農林水産省2011,河田ら 2000,中矢ら 2007,など)を参照されたい.
(2)栽培管理
水稲では活着期に高塩条件に対する感受性が高い.したがって,塩害水田では出来る限り若い苗の移植は避けた方が好ましい.また,苗の活着を高めるため,強風や低温時の移植は控え,移植時の水管理は深めがよいとされている.また移植後は用水の掛け流しや,湛水することにより土壌塩類濃度の上昇を抑制することが望ましい.塩害によって分げつ数の抑制が予想される場合には,植え付け本数や栽植密度の増加による穂数確保が推奨される.また,移植直後は塩害を受けやすいので,除草剤は活着を確認した後に散布する.
水管理においては,落水状態では土壌中の塩類の表層への蓄積が促進されることに留意する必要がある.除塩が不十分な水田で干天が続くような場合には,中干しの中止や中干し期間の短縮を行う.特に穂ばらみ期および出穂開花期の高塩条件は不稔を生じるので留意が必要である.
東日本大震災では用水路に海水が流れ込んだり,液状化により地下からの塩類が用水に混入した事例がみられる.用水に塩類が含まれる場合には,乾燥ストレスと塩ストレスの両方を軽減する必要があるため水管理には困難が伴う.灌水による圃場への塩類の流入,蓄積を軽減するためには節水栽培が必要である.一方で土壌が乾燥しすぎるとイネが水ストレスを受けたり,塩類が土壌表層に集積しやすい.農業用水基準(水稲)では,用水の塩類濃度は0.3 dS/m以下とされている(農林水産省 1970年).北海道で用水に海水が混入した事例では,塩分濃度0.1%(EC2.0dS/m)を超える用水は圃場を乾燥させないような入水にとどめ,活着後~幼穂形成期では0.25%(EC4.5dS/m),幼穂形成期以降では0.20%(EC3.7dS/m)以上で取水を停止することを推奨している(北海道立中央農業試験場2001).IRRIでは灌漑水について以下のような目安を示している(IRRI);
・pH6.5~8.0, EC<0.5 dS/m 灌漑に適している
・pH8.0~8.4, EC 0.5~2.0 dS/m 中程度の質の灌漑水である
・pH>8.4, EC>2.0 dS/m 灌漑には適さない
実際には,用水の塩類濃度をモニタリングしながら,生育ステージや土壌のECや水分に応じて水管理を行うことが望ましい.
畑条件かつハウス内で栽培する花き等では,培地中の水は栽培で灌水に使った水に比べ塩類が濃縮される.そのため,栽培に使う水の塩類濃度に注意するとともに,塩類濃度の高い用水を使った場合は,栽培後培地の除塩を行うのが望ましい.
(3)施肥管理
水稲では,土壌が貧栄養の条件下では生育が低下するだけではなく,高塩による被害程度も拡大することが報告されている(曽根ら 2011).そのような貧栄養かつ高塩の条件下では,生育の向上に窒素施肥が最も効果的であり,次にリン施肥によって収量が向上した(曽根ら 2009).高塩条件下で適度な窒素施肥が塩害を軽減することは,コムギ,トウモロコシ等でも報告されている.しかしながら,高塩類条件では窒素過剰害が起こりやすいことや(Yamamotoら 2004),倒伏を助長するため,過剰な施用は控える.また塩害は土壌還元害を引き起こすことがあるため,有機物の資材の施用には留意する.畑条件では硝酸化成過程は塩類の影響を受けやすい(池田ら1987).畑栽培では尿素や硫安より硝安などの硝酸態窒素が望ましい.また,土壌pHが上昇した場合,可給態リンが減少しリン欠乏状態になりやすい.
液状化による噴砂が残っているような圃場では,ECや可給態窒素,リン酸に圃場ムラが生じている可能性もある.したがって,栽培を行う前に土壌EC値等で圃場ムラを調査し,基肥についてはリン酸を増やし,生育に応じて分げつ期以降に窒素を追肥することが望ましい.
石膏(CaSO4)など石灰資材はしばしば塩害軽減の土壌資材として用いられる.石灰資材による生育促進は,Caによる土壌の物理化学性の改良と植物体のCaとNaのバランスの改善の双方によると考えられる.高NaCl条件でCaを添加するとイネのNa吸収・地上部への移行は抑制され,生育阻害が緩和された(Anil ら 2005,山内ら 1983).ただし水田ではCaSO4など硫酸根をもつ資材の投入が,H2S発生による生育阻害を増大する可能性もあるので留意が必要である.
またケイ酸は,耐塩性を向上させるが,その要因としてはNa吸収の抑制や葉の蒸散抑制などが寄与していると考えられる(Matoh ら 1986).イネの根ではバイパスフローによりNaの流入があるが,根でのケイ酸の沈着が塩ストレス下でのNa吸収を抑制する(Gong et al. 2006).土壌の可給態ケイ酸が低い場合には施肥効果が期待される.
Naと同じ1価の陽イオンであるKは,多くの代謝過程においてNaと拮抗関係にある.このためK施用により生育抑制がNa害が軽減された例もある(Bohra and Doerffling 1993).しかし,Kの増肥は総イオン濃度の上昇にもつながるため過剰な施用は控えるのが望ましい.
おわりに
被災された多数の皆様に対して,心からお見舞いを申し上げますと共に,農業農村の1日も早い復興を祈念いたします.
引用文献
- Ahmadi N, Negrao S, Katsantonis D, Frouin J, Ploux J, Letourmy P, Droc G, Babo P, Trindade H, Bruschi G, Greco R, Oliveira MM, Piffanelli P, Courtois B.(2011)Targeted association analysis identified japonica rice varieties achieving Na+/K+homeostasis without the allelic make-up of the salt tolerant indica variety Nona Bokra. Theor. Appl. Genet. DOI 10.1007/s0122-1634-4
- Akbar M, Yabuno T, Nakao S.(1972)Breeding for saline-resistant varieties of rice I. Variability for salt tolerance among some rice varieties. Jap. J. Breed. 22:277-284
- Akita S, Cabuslau GS.(1990)Physiological basis of differential response to salinity in rice cultivars. Plant and Soil 123:277-295.
- Anil VS, Krishnamurthy P, Kuruvilla S, Sucharitha K, Thomas G, Mathew MK.(2005)Regulation of the uptake and distribution of Na+in shoots of rice(Oryza sativa)variety Pokkali:role of Ca2+in salt tolerance response. Physiologia Plamtarum.124:451-464.
- Bohra JS. And Doerffling K.(1993)Potassium nutrition of rice(Oryza sativa L.)varieties under NaCl salinity. Plant and Soil 152:299-303.
- Chandrasekharan, H., Sarangi, A., Nagarajan, M., Singh, V. P., Rao, D. U. M., Stalin, P., Natarajan, K., Chandrasekaran, B. and Anbazhagan, S.(2008)Variability of soil-water quality due to Tsunami-2004 in the coastal belt of Nagapattinam district, Tamilnadu. Journal of Environmental Management, 89:63-72.
- Dasgan HY, Aktas H, Abak K, Cakmak I.(2002)Determination of screening techniques to salinity tolerance in tomatoes and investigation of genotype responses. Plant Sci. 163:685-703.
- Datta, S.K. 1972. A study of salt tolerance of twelve varieties of rice. Curr. Sci. 41:456-457.
- Flowers TJ and Yeo AR(1981)Variability in the resistance of sodium chloride salinity within rice(Oryza sativa)varieties. New Phytol. 88:363-373.
- Flowers TJ, Hajibagheri MA and Yeo AR.(1991)Ion accumulation in the cell walls of rice plants growing under saline conditions;evidence for the Oertli hypothesis. Plant cell environ. 14:319-325.
- Genc Y, Oldach K, Verbyla AP, Lott G, Hassan M, Tester M, Wallwork H, McDonald GK.(2010)Sodium exclusion QTL associated with improved seedling growth in bread wheat under salinity stress. Theor. Appl. Genet. 121:877-894
- Golldack D, Quigley F, Michalowski CB, Kamasani UR, Bohnert HJ(2003)Salinity stress-tolerant and -sensitive rice(Oryza sativa L.)regulate AKT1-type potassium channel transcripts differently. Plant Mol. Biol.351:71-81
- Gong HJ, Randall DP, Flowers TJ(2006)Silicon deposition in the root reduces sodium uptake in rice(Oryza sativa L.)seedlings by reducing bypass flow. Plant cell environ. 29:1970-1979
- Gorham J, Jones RGW, Bristol A(1990)Partial characterization of the trait for enhanced K+-Na+discrimination in the D genome of wheat. Planta 180:590-597.
- IRRI Rice Knowledge Bank, Rice Doctor, Salinity http://www.knowledgebank.irri.org/RiceDoctor/index.php?option=com_content&view=article&id=548&Itemid=2753
- James RA, Davenport RJ, Munns R(2006a)Physiological characterisation of two genes for Na+exclusion in durum wheat:Nax1 and Nax2. Plant Physiol. 142:1537-1547
- James RA, Munns R, Caemmerer von S, Trejo C, Miller C, Condon T.(2006b)Photosynthetic capacity is related to the cellular and subcellular partitioning of Na+, K+and Cl- in salt-affected barley and durum wheat. Plant cell environ. 29:2185-2197.
- Kume, T., Umetsu, C. and Palanisami, K.(2009)Impact of the December 2004 tsunami on soil, groundwater and vegetation in the Nagapattinam district, India. Journal of Environmental Management, 90:3147-3154.
- Leidi EO and Saiz JF(1997)Is salinity tolerance related to Na accumulation in upland cotton(Gossypium hirsutum)seedlings?Plant Soil 190:67-75
- Lin, H. X., M. Z. Zhu, M. Yano, J. P. Gao, Z. W. Liang, W. A. Su, X. H. Hu, Z. H. Ren, D. Y. Chao 2004. QTLs for Na+and K+uptake of the shoots and roots controlling rice salt tolerance. Theor. Appl. Genet. 108:253-260
- Lutts L, Kinet JM, Bouharmont J(1996)Effects of salt stress on growth, mineral nutrition and proline accumulation in relation to osmotic adjustment in rice(Oryza sativa L.)cultivars differing in salinity resistance. Plant Growth Regulation 19:207-218
- Maas and Hoffman(1977)Crop salt tolerance-current assessment J. Irr. and Drainage Div. ASCE 103:115-134
- Marschner H(1995)Saline soil. In Mineral nutrition of higher plants. Academic press. London UK, pp657-680.
- Matoh T, Kairysmee P, Takahashi E(1986)Salt-induced damage to rice plants and alleviation effect of silicate. Soil Sci Plant Nutri. 32:295-304
- Matsushita N and Matoh T(1992)Function of the shoot base of salt-tolerant reed(Pharagmites communis Trinius)plants for Na+exclusion from the shoots. Soil Sci Plant Nutri. 38:565-571.
- Moradi F and Ismail AM(2007)Responses of photosynthesis, chlorophyll fluorescence and ROS-scavenging system to salt stress during seedling and reproductive stages in rice. Annals. Bot. 99:1161-1173.
- Munns R and James R(2003)Screening methods for salinity tolerance:a case study with tetraploid wheat. Plant Soil 253:201-218
- Munns R and Tester M(2008)Mechanisms of salinity tolerance. Ann. Rev. Plant Biol. 59:651-681
- Nakaya T, Tanji H, Kiri H and Hamada H.(2010)Developing a salt-removal plan to remedy tsunami-caused salinity damage to farmlands:Case study for an area in southern Thailand. JARQ, 44:159-165.
- Raja R, Chaudhuri SG, Ravisankar N, Swarnam TP, Jayakumar V and Srivastava RC(2009)Salinity status of tsunami-affected soil and water resources of South Andaman, India. Curr. Sci. 96:152-156.
- Ren Z-H, Gao J-P, Li L-G, Gai X-L, Huang W, Chao D-Y, Zhu M-Z, Wang Z-Y, Luan S, Lin H-X.(2005). A rice quantitative trait locus for salt tolerance encodes a sodium transporter. Nature Genetics 37:1141-1146.
- Senadhira D, Zapata-arias FJ, Gregorio GB, Alejar MS, Cruz de la HC, Padolina TF, Galvez AM.(2002)Development of the first rice cultivar through indica/japonica anther culture. Field Crops Res. 76:103-110
- Thomson, M. J., M. de Ocampo, J. Egdane, M. A. Rahman, A. G. Sajise, D. L. Adorada, E. Tumimbang-Raiz, E. Blumwald, Z. I. Seraj, R. K. Singh, G. B. Gregorio, A. M. Ismail(2010)Characterizing the saltol quantitative trait locus for salinity tolerance in rice. Rice 3:148-160.
- US Soil Survey Staff(1951)Estimation and mapping of salt and alkali on the soil. Soil survey manual. US Dept Agric. Handbook.
- Walia H, Wilson C, Condamine P, Liu X, Ismail AM, Zeng L, Wanamaker SI, Mandal J, Xu J, Cui X, Close TJ(2005)Comparative transcriptional profiling of two contrasting rice genotypes under salinity stress during the vegetative stage. Plant Phsyiol. 139:822-835
- Walia H, Wilson C, Zeng L, Ismail AM, Condamine P, Close TJ(2007)Genome-wide transcriptional analysis of salinity stresses japonica and indica genotypes during panicle initiation stage. Plant Mol. Biol. 63:609-623.
- Yamamoto A, Shim IS, Fujihara S, Yoneyama T and Usui K(2004)Effect of difference in nitrogen media on salt-stress response and contents of nitrogen compounds in rice seedlings. Soil Sci. Plant Nutri. 50:85-93.
- Yeo AR and Flowers TJ(1986)Salinity resistance in rice(Oryza sativa L.)and a pyramiding approach to breeding varieties for saline soils. Aust. J. Plant Physiol. 13:161-173.
- Yeo AR, Caporn SJM, Floweres TJ(1985)The effect of salinity upon photosynthesis in rice(Oryza sativa L). I:gas exchange by individual leaves in relation to their salt content. J.Exp. Bot. 36:1240-1248.
- Yeo AR, Yeo ME, Flowers SA, Flowers TJ(1990)Screening of rice(Oryza sativa L.)genotypes for physiological characters contributing to salinity resistance, and their relationship to overall performance. Theor. Appl. Genet. 79:377-384.
http://www.pref.saga.lg.jp/web/shigoto/_1075/_32933/ns-nousisetu/boujocenter/_10660/kisyougai/enngai/enngaiyasai.html - 愛知県 1963 愛知県農業試験場彙報 18号
- 加藤直樹,佐藤健次,服部育男,山本克己,原口暢朗,久保田哲史(2006)干陸地土壌での飼料イネの初期生育の品種比較. 日本草地学会誌 52(別2):188-189.
- 河田直美・有好利典・大内英司(2000)平成11年台風18号による塩害対策―38年ぶりの実施―. 水と土 121:10-17.
- 宮城県立農業試験場(1961)チリ沖地震津波による農作災害の実態74pp
- 近藤始彦・荒井(三王)裕見子・小林伸哉・高井俊之・岩澤紀生・吉永悟志(2011)塩ストレスの水稲品種生育への影響比較日本作物学会関東支部会報 26:28-29
- 高橋栄一 1987. 生命にとって塩とは何か 土と食の塩過剰 農文協.
- 佐賀県(2008a).塩害対策野菜.
- 佐賀県(2008b).塩害対策水稲.http://www.pref.saga.lg.jp/web/shigoto/_1075/_32933/ns-nousisetu/boujocenter/_10660/kisyougai/enngai/enngaisui.html
- 山内益夫・前田吉広・長井武雄(1987)耐塩性の品種間差とナトリウムの吸収・移行特性との関係(1)土肥誌58:591-594.
- 山内益夫・島田義明・吉田昌一(1983)水稲における塩化ナトリウム高濃度処理による生育障害の軽減に対するカルシウムイオンの効果. 土肥誌54:499-504.
- 曽根千晴・坂東正章・津田誠・平井儀彦(2009)N,PおよびKの施与が塩条件下のNERICA 1の乾物生産および栄養素吸収に及ぼす影響.日作紀79別(2):148―149.
- 曽根千晴・津田誠・平井儀彦(2011)土壌の違いが種間雑種NERICA系統の塩条件下におけるNa蓄積におよぼす影響.日作紀 80:333―340.
- 池田順一・小林達治・高橋英一(1987)共生窒素固定および土壌中でのアンモニア化成・硝酸化成に及ぼす塩類ストレスの影響 土肥誌 58:53-57
- 中矢哲朗・丹治肇・桐博英・濱田浩正(2007)津波による農地塩害軽減のための除塩計画策定手法. 土木学会 海岸工学論文集 54:1386-1390.
- 中矢哲郎・丹治肇・桐博英(2006)2004年インド洋津波によるタイ南部農村地帯の長期的被害調査 海岸工学論文集53:1381-1385
- 嶋田典司(1972)千葉大学園芸学部特別報告,6
- 農林水産省(2011)農地の除塩マニュアル;農村振興局http://www.maff.go.jp/j/press/nousin/sekkei/pdf/110624-01.pdf
- 北海道立中央農業試験場(2001)塩分を混入するかんがい用水の取水管理. 平成13年度研究成果情報 北海道農業
- 木田義信・佐々木園子・佐藤正一(2007)土壌塩分が水稲苗の活着に及ぼす影響. 東北農業研究. 60:35-36
寄稿
近藤始彦1・曽根千晴2・荒井(三王)裕見子1・小林伸哉1・高井俊之1・岩澤紀生1・吉永悟志1
1.農研機構・作物研究所,2.国際農林水産業研究センター